








SUMO化及びO-GlcNAc化によるMAPキナーゼ経路の活性制御機構と疾患
 |
研究代表者 |
 |
研究分担者 冨田 太一郎 東京大学 医科学研究所 分子細胞情報分野 |
【研究概要】
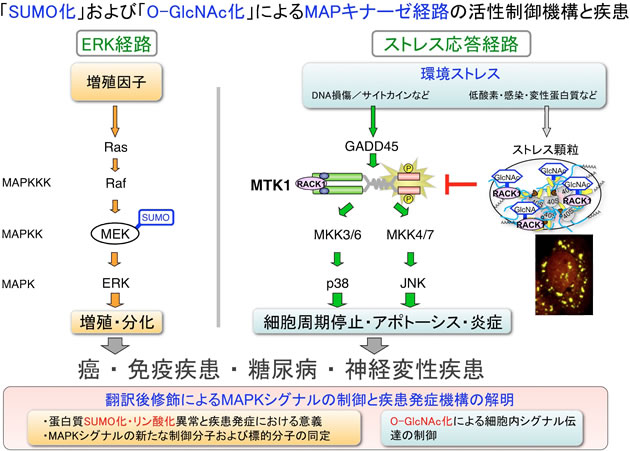
ヒト細胞内には、主に増殖因子によって活性化され、細胞増殖や生存に作用するERK経路と、様々な環境ストレス刺激に応答して活性化され、細胞死(アポトーシス)や炎症を惹起するストレス応答MAPキナーゼ(p38及びJNK)経路という、複数のMAPキナーゼ・カスケードが存在する。細胞運命(生か死か)を決定して生体の恒常性維持を担うこれらMAPK経路の制御異常が、癌や自己免疫疾患などの発症に深く関与することが明らかにされている。しかしながら、MAPキナーゼ経路の活性調節機構や疾患における制御異常の詳細には、未だ不明な点が数多く残されており、その解明は疾患克服の観点からも重要である。
私達は最近、ERK経路のMAPKKであるMEKが、細胞内でSUMO化されること、さらにその結果、MEK活性が阻害されてERK経路が負に制御されることを見出した。即ち、MEKのSUMO化はERK経路の過剰な活性化を防ぎ、増殖シグナル強度の適切な制御に重要な役割を果たしていることを明らかにした。さらに私達は、癌遺伝子である活性型RasがMEKのSUMO化を阻害することを見出し、実際にRasに変異を持つ癌細胞内においてMEKのSUMO化が消失していることを明らかにした。従って、癌遺伝子RasはRafを活性化すると同時に、MEKのSUMO化による不活性化を阻止する、という二重の機構によってERK経路を強く活性化し、発癌を導くと考えられる。本研究では、MEKのSUMO化異常が発癌に果たす役割をより詳細に解析すると共に、SUMO化やリン酸化などの翻訳後修飾によるMAPKシグナルと生命機能の制御メカニズムを分子レベル・個体レベルで解明する。さらにMAPK経路の新たな活性制御分子や未知の標的分子を同定して、癌や自己免疫疾患を代表とする難治性疾患の病因・病態形成との関連を明らかにする。
また一方で私達は、蛋白質のO-GlcNAc化修飾がMAPKシグナルの活性制御やストレス顆粒(細胞のストレス適応を担う構造体)の形成に関与することを示す知見を得ている。本研究においては、O-GlcNAc化が細胞内シグナル伝達の制御に果たす役割を解明すると共に、その異常と疾患発症との関連を検証する。また、構造生物学、数理科学や生体イメージングの研究者と連携して、SUMO化、リン酸化、およびO-GlcNAc化によるMAPKシグナル伝達ネットワークの時空間制御の全容を理解し、疾患に対する新たな分子診断法や治療法開発への応用・発展を目指す。
【参考文献】
- Nakamura T, Saito H and Takekawa M. SAPK pathways and p53 cooperatively regulate PLK4 activity and centrosome integrity under stress. Nature Commun. 4:1775 doi: 10.1038/ncomms2752 (2013)
- Tomida T, Osa S, Takekawa M, Iino Y and Saito H. The temporal pattern of stimulation determines the extent and duration of MAPK activation in a caenorhabditis elegans sensory neuron. Science Signaling 5, ra76 (2012)
- Kubota Y, O’Grady P, Saito H and Takekawa M. Oncogenic Ras abrogates MEK SUMOylation that suppresses the ERK pathway and cell transformation. Nature Cell Biol. 13, 282-291 (2011)
- Tomida T, Takekawa M, O’Grady P and Saito H. Stimulus-specific distinctions in spatial and temporal dynamics of stress-activated protein kinase kinase kinases revealed by FRET biosensor. Mol. Cell. Biol. 29, 6117-6127 (2009)
- Arimoto K, Fukuda H, Imajoh-Ohmi S, Saito H, and Takekawa M. Formation of stress granules inhibits apoptosis by suppressing stress-responsive MAPK pathways. Nature Cell Biol. 10: 1324-1332 (2008)
- Miyake Z, Takekawa M, Ge Q and Saito H. Activation of MTK1/MEKK4 by GADD45 through induced N-C dissociation and dimerization-mediated trans-autophosphorylation of the MTK1 kinase domain. Mol. Cell. Biol. 27, 2765-2776 (2007)
- Nakajima A, Komazawa-Sakon S, Takekawa M, Sasazuki T, Yeh W-C, Yagita H, Okumura K and Nakano H. An antiapoptotic protein, c-FLIPL, directly binds to MKK7 and inhibits the JNK pathway. EMBO J. 25, 5549–5559 (2006)
- Takekawa, M., Tatebayashi, K., and Saito, H. Conserved docking site is essential for activation of mammalian MAP kinase kinases by specific MAP kinase kinase kinases. Molecular Cell 18, 295-306 (2005)
- Takekawa, M., Tatebayashi, K., Itoh, F., Adachi, M., Imai, K. and Saito, H. Smad-dependent GADD45β expression mediates delayed activation of p38 MAP kinase by TGF-β. EMBO J. 21, 6473-6482 (2002)
- Takekawa, M. and Saito, H. A family of stress-inducible GADD45-like proteins mediate activation of the stress-responsive MTK1/MEKK4 MAPKKK. Cell 95, 521-530, (1998)



